초록
동결 건조된 유산균의 저장 안정성은 비용 효율성에 중요한 요소이다. 유산균의 장기 보관은 미생물 산업이 유통 비용을 절감할 수 있도록 한다. 여기서는 동결 건조 과정과 후속 저장 과정에서 류코노스톡 메센테로이드 WiKim32의 생존 가능성에 대한 -5°C의 과냉각 조건에서 냉적응의 영향을 조사했다. 저온 적응은 외다당류(EPS)의 두께를 증가시키고 동결 건조된 Leu. mesenteroides WiKim32의 실행 가능성을 향상시켰다. 저온 적응 셀은 비적응 셀에 비해 과냉각 조건에서 EPS 두께가 35.4% 증가하는 것으로 나타났다. EPS 가수분해 세포의 생존 가능성은 처리되지 않은 세포보다 낮았으며, 이는 EPS가 동결 건조 과정에서 보호에 역할을 한다는 것을 의미한다. 저온 적응은 동결 건조된 Leu. mesenteroide WiKim32의 저장 안정성을 증가시켰다. 보관 후 56일이 지난 시점에서 -5°C의 저온 적응을 통해 가장 높은 생존율(71.3%)을 달성했습니다. 동결건조 전 EPS 함유 육수를 첨가했을 때 생존율은 82.7%로 더욱 높아졌다. 이러한 결과는 과냉각 전처리에 의한 저온 적응이 Leu. mesenteroides WiKim32의 장기 저장을 위한 좋은 전략이 될 것임을 시사한다.
상용화된 LAB 스타트업은 공식화 중에 매우 실행 가능하고 후속 저장 중에 매우 안정적이어야 한다. 동결 건조 및 동결 건조 방법은 일반적으로 LAB 시동기의 세포 생존성 및 저장 안정성을 극대화하기 위해 사용된다. 그러나 이러한 방법들은 LAB 시작자들이 공정 중에 얼음 결정 형성 및 삼투압 응력과 같은 불리한 조건에 직면하게 하여 세포막 손상을 초래한다[5].
저온 조건으로 인한 부정적인 영향을 제한하기 위해 다양한 저온 보호제가 도입되고 있다. 이러한 에이전트는 동결 건조 프로세스 전에 LAB 시동기와 혼합되어 실행 가능성을 보장합니다[6]. 이러한 작용제는 분자량에 따라 저분자량 작용제와 고분자량 작용제의 두 가지 유형으로 분류된다. 종래의 것들, 즉 젖당, 수크로스 및 트레할로스는, 변성 및 낮은 수분 함량에 의한 손상을 방지하여, 세포막의 극성기 및 단백질과 수소 결합을 형성한다[7]. 후자, 즉 탈지 우유, 말토덱스트린 및 콩 분말은 독성, 가용성 및 생분해성 때문에 주로 LAB 보호에 사용된다[8, 9].
LAB는 생리학적 스트레스, 즉 동결 건조 과정 전의 저온 조건에 대한 노출에 반응하여 동결 내성을 개발할 수 있다. LAB 성장에 필요한 최적 온도 미만의 온도는 LAB 실행 가능성을 향상시키는 콜드 쇼크 반응의 시작을 초래한다. 이러한 반응에는 감기 관련 유전자의 상향 조절, 감기 유도 단백질의 생성, 지방산 조성의 변화, 양이온 유입의 증가가 포함된다[10-13]. 저온 적응 세포는 동결 건조 과정 후에 결국 살아남는다. 상기 연구들은 락토바실러스(Lactobacillus spp.)에 초점을 맞추고 있지만, 동결건조 후 다른 속의 LAB를 장기간 보관하는 것이 최근에 더욱 주목받고 있다.
김치, 파오카이, 사우어크라우트와 같은 발효 채소 음식의 감각적 특성은 Leuconostoc spp를 사용하여 강화될 수 있다. 발효 초기에 김치로부터 격리된 Leu. mesenteroide WiKim32는 특히 김치의 유통기한을 연장하고, 김치의 감각적 특성을 향상시키는 장점으로 인해 김치 스타터로 사용되어 왔다[14]. 이전 연구에 따르면 -5°C에서 과냉각 전처리를 수행하면 Lac. brevis WiKim0069의 저장 안정성이 향상됩니다[13]. 본 연구의 전반적인 목적은 Leu. mesenteroides WiKim32의 생존력을 향상시키는 외다당류(EPS) 생성 및 세포벽 조성에 대한 냉적응의 영향을 조사하는 것이었다. 또한 동결건조 전 다양한 농도의 EPS를 첨가하는 것이 세포생존성에 미치는 영향을 조사하였다.
세균주 및 배양조건
김치에서 분리되어 김치 스타터로 개발된 Leu. mesenteroides WiKim32(KFCC11639P)는 세계김치연구소(광주광역시)의 배양 컬렉션에서 얻은 것이다. 주식 배양액은 데만, 로고사, 샤프(MRS) 배지(미국 Difco Laboratories)의 -80°C(MDF4V, 일본 Panasonic)에 글리세롤 20%로 저장하였다. 작업 배양액은 MRS 배지에서 30°C에서 24시간 동안 배양되었다.
냉간 적응 절차 및 셀 실행 가능성
저온 적응 과정과 세포 생존력 측정은 최 등이 설명한 절차를 이용하여 수행하였다. [13. Leu. mesenteroide WiKim32는 30°C에서 24시간 MRS 배지에서 배양하고 원심분리(25°C에서 20분 동안 10,000 × g)를 통해 수확하였다. 여분의 MRS 육수는 후속 실험을 위한 현탁액으로 4°C에서 저장되었다. 원심분리를 통해 수집된 침전된 세포는 멸균 인산염 완충 식염수(3M Company, USA)로 2회 세척하고 식염수 또는 24h 배양 MRS brother에 4 × 1010 CFU/ml로 농축하였다. 최종 세포 농도는 동결방지제로서 콩가루 20% 용액의 균일한 부피로 2 × 1010 CFU/ml로 조정되었다. 셀들은 동결 건조 전에 10°C 또는 -5°C에서 2시간 동안 차가운 응력을 받았다. 대조 검체는 저온 스트레스에 노출되지 않았습니다. 동결건조 공정은 동결건조기(FDCF-12012; Operon, 대한민국)를 사용하여 진공도가 1Pa 미만인 -80°C에서 실시하였다. 동결건조된 셀은 보관기간별로 별도로 준비하여 -20°C에서 56일간 보관하였다.
동결건조 직후와 저장기간 동안 판수법을 이용하여 세포의 생존가능성을 평가하였다. 적절한 희석제를 MRS 한천에 붓고 30°C에서 48시간 동안 배양하였으며, 세균군락은 CFU/ml로 계산하였다.
EPS 및 EPS 프리 박테리아의 제조
EPS 정화. 최씨 등이 보고한 절차에 따라 EPS를 정제했다. [15], 약간의 조정이 필요합니다. 세포수확 후, 불활성화효소를 위해 100°C에서 5분간 폐기된 MRS배지를 끓인 후 원심분리(4°C에서 20분간 10,000 ×g)를 통해 상등액을 얻었다. 에탄올 침전 및 동결건조를 위해 상등액에 95% 에탄올의 3배를 첨가하여 원심분리 전 -20°C에서 밤새 유지하였다(4°C에서 10,000 ×g은 20분간 20분간). 단백질은 증류수에 용해시킨 후 4% 트리클로로아세트산으로 탈단백질화하여 제거하였다(Sigma-Aldrich, USA). 원유 EPS를 95% 에탄올로 재침하하고 증류수에 용해시킨 후 Slide-A-Lyzer 투석 카세트(10K MWCO, Thermo Scientific, USA)를 사용하여 4°C에서 2일간 증류수로 투석하고 -20°C에서 하룻밤 동안 건조하였다. 정제된 EPS는 동결방지제로서 세포생존성 시험에 첨가되었다.
EPS의 효소 가수분해. Leu. mesenteroides Wikim32의 EPS 함량을 줄이기 위해 세균세포를 가수분해효소인 Dextranase Plus L(Novozyme, Chaetomium rancelicum)로 처리하였다. 이어서 세균배양배지 200ml에 덱스트라나제 용액 500μl를 첨가하였다. 효소 가수분해는 30°C에서 12시간 동안 수행되었다. 상기 EPS 가수분해 셀은 10,000 × g에서 4°C에서 20분간 원심분리하여 수확한 후 증류수로 3회 세척하였다. EPS 가수분해 세포의 생존 가능성은 판수법을 이용하여 0.1% 정제된 EPS를 동결보호제로 사용하거나 사용하지 않고 측정하였다.
3. 결과
Leu. mesenteroides WiKim32의 EPS 두께에 대한 냉간 적응의 영향
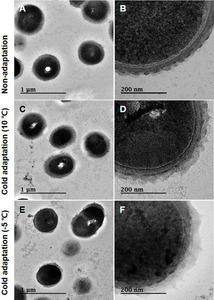
박테리아 세포벽의 구조적 변화는 투과전자현미경(TEM)을 이용하여 측정하였다(그림 1). Leu. mesenteroides WiKim32의 EPS 두께는 전처리 조건(F = 62.4, df = 2,45, p < 0.001)에 따라 변화하였다. 가장 두꺼운 EPS(51.6 ± 3.52 nm)는 -5°C의 저온 적응 셀에서 관찰되었으며, 이어서 10°C(42.0 ± 3.34 nm; 표 1)의 저온 적응 셀에서 관찰되었다. 저온 적응 세포의 EPS 두께는 비적응 세포에 비해 10.2~35.4% 증가하였다. 단당 조성은 EPS 두께가 적응된 조건에 의존하는 이유를 강력하게 뒷받침했다. 비적응 세포의 탄수화물은 주로 아라비노스(1.6%), 갈락토스(2.8%), 포도당(8.7%)으로 구성된다(표 2). 세포가 -5°C에서 적응된 후 탄수화물 함량은 아라비노스(1.7%), 갈락토스(2.9%), 포도당(9.6%)으로 변화했다. LAB의 세포벽에는 (a) 외층, 펩티도글리칸 및 리포테이초산으로 구성된 세포벽, (b) EPS로 둘러싸인 세포벽, (c) 표면층 단백질(SLP)로 둘러싸인 세포벽, (d) 세포벽 가교된 EPS 및 SLP[13]의 네 가지 구조 모델이 있다. 이들 모델 중 Leu. mesenteroides WiKim32는 EPS가 세포벽에 부착된 세포벽 모델을 가지고 있다[15]. 본 연구에서는 과냉각 전처리를 통해 Leu. mesenteroides WiKim32의 EPS 두께를 더욱 증가시켰다.
EPS의 효소적 가수분해가 세포생존성에 미치는 영향
Leu. mesenteroides WiKim32의 생존성에 대한 EPS의 역할을 이해하기 위해 EPS 가수분해 세포를 제조하였다(그림 2). 세포벽에 부착된 EPS의 가수분해에 의해 포도당 함량이 감소하였으며, 이는 EPS가 성공적으로 제거되었음을 시사하였다(표 2). 세포 유형(F = 173.0, df = 1,12, p < 0.001)에서 유의미한 생존율 차이가 관찰되었다(그림 3). EPS 가수분해 세포의 생존 가능성은 처리되지 않은 세포보다 낮았으며, 이는 EPS가 동결 건조 과정에서 보호에 역할을 한다는 것을 시사한다. LAB의 생존 가능성은 EPS와 콩가루를 포함한 보호제에 의해 심각하게 영향을 받는다. 세포의 종류에 관계없이 보호제가 없는 경우(F = 2927.5, df = 2,12, p < 0.001) 가장 낮은 생존율을 기록하였다. EPS를 보호제로 사용한 결과, 치료되지 않은 세포와 EPS 가수분해된 세포 모두에서 각각 18.6%, 12.1%의 생존율을 보였다. 콩가루를 보호제로 사용한 미처리 세포에서 89.6%로 가장 높은 생존율을 보였고, 콩가루를 사용한 EPS 가수분해 세포에서 66%로 뒤를 이었다.
미생물 EPS는 세포 표면에 영구적으로 부착되지 않은 세포 외 탄수화물 다당류이다. LAB에서 생산된 EPS는 항암, 항산화, 항바이러스, 면역자극제 및 콜레스테롤 강하 효과와 같은 다양한 건강 유익 특성을 보여주었다[15, 16]. 그들은 또한 탈수, 불리한 온도, 삼투압 스트레스 및 항생제와 같은 환경 스트레스로부터 Leu. mesenteroides WiKim32를 보호하는 것을 돕는다[17]. 본 연구에서는 EPS의 제거로 인해 낮은 생존성이 나타났으며, 이는 세포 표면을 둘러싸는 EPS 층이 동결 건조 과정에 대한 보호 활성을 나타냄을 보여준다. 동결 건조 전에 몇 개의 EPS를 추가하면 Leu. mesenteroides WiKim32의 실행 가능성이 향상된다는 점을 고려하면, 더욱이 EPS는 보호제로 사용될 수 있다.
과냉각 전처리에 의한 냉간적응이 안정성에 미치는 영향
동결건조 Leu. mesenteroide WiKim32 셀의 저장 안정성은 전처리 조건(F = 166.7, df = 2,24, p < 0.001), 저장 시간(F = 60.4, df = 1,24, p < 0.001), 현탁액의 종류(F = 65.6, df = 1,24, p < 0.001)에 따라 변화하였다(그림 4). 냉동 건조 및 후속 저장 시 저온 적응 세포는 비적응 세포보다 생존력이 높았다. 동결건조 후 저온적응세포의 생존율은 80% 이상을 유지한 반면 비적응세포는 67.2%로 생존율이 크게 감소했다. 전처리 방법 중에서는 -5°C에서의 냉적응이 28일 보관 후 86.8%를 보여 생존력 향상에 가장 우수했다. 동결건조된 Leu. mesenteroides WiKim32 세포의 생존가능성은 멸균식염수와 24시간 배양된 MRS배지를 포함한 현탁액의 유형과 달랐다. EPS가 0.1%(w/v) 함유된 MRS배지에서 배양된 Leu. mesenteroides WiKim32 세포의 안정성은 식염수보다 높았다. 저온 적응 Leu. mesenteroides WiKim32의 24시간 배양 MRS 육수에서 -5°C에서 최대 56일간 생존율 82.7%가 유지되었고, 이어서 10°C에서 저온 적응 세포가 사용되었다.
이전 연구에서, 저온 적응은 동결 건조 락토바실러스 브레비스 WiKim0069의 저장 수명 연장의 이점을 가지고 있다[12, 13]. 빙점 이하의 과냉각 조건은 동결 건조에 대한 Lac. brevis WiKim0069의 저항을 더욱 증가시켜 10°C에서의 저온 적응보다 더 높은 생존력으로 이어졌다[13]. 세포가 적응 조건에 노출되면 양이온 유입과 SLP 생성이 모두 증가한다. Ca2+ 및 Mg2+와 같은 세포 내 양이온은 칼슘 의존성 세린-트레오닌 포스파타아제로 알려진 칼시뉴린의 활성화 및 삼투압 스트레스에 대한 내성을 유도할 수 있다[18-21]. 저온 적응으로 인한 SLP 형성은 동결 건조로부터 세포를 보호하기 위해 세포 표면을 두껍게 만들었다. 표면층 두께는 저온 적응 조건에 따라 결정된다. 마찬가지로, EPS는 세포의 접착에 중요한 역할을 하며 세포에 보호 장벽을 제공한다. EPS가 0.1%(w/v) 존재할 때 Leu. mesenteroides WiKim32 세포의 생존율은 10% 이상 증가했다. 우리의 결과는 김 외 연구진의 이전 연구와 일치한다. [22], Pseudoaltermonaselyakovii에 의해 생성된 EPS의 존재가 동결-퇴출 주기에 대한 대장균의 내성을 향상시켰음을 나타낸다. EPS의 존재는 Leu. mesenteroides WiKim32가 차가운 스트레스에 저항하는 것을 도와주며, 이는 이것이 저온 보호제로 사용될 수 있음을 시사한다. 과냉각 전처리에 의한 저온 적응은 류코노스톡 spp. 및 락토바실러스 spp.와 같은 유산균의 장기 저장을 위한 좋은 전략이 될 것이다.



